manbet手机版先进的信息
manbet手机版科学背景:
manbet手机版关于灭绝古人类基因组和人类进化的发现(pdf)
manbet手机版科学背景
manbet手机版关于已灭绝古人类基因组和人类进化的发现
manbet手机版Svante Pääbo获得了2022年诺贝尔生理学或医学奖,以表彰他在灭绝古人类基因组和人类进化方面的发现。manbet手机版智人与已灭绝的古人类之间的关系一直是人们非常感兴趣的话题。manbet手机版古生物学和考古学对人类进化的研究很重要。manbet手机版现代DNA技术为我们更精确地研究远古的过去提供了机会。manbet手机版然而,由于DNA在数万年间的降解以及微生物和当代人类的污染所带来的极端技术挑战,长期以来人们一直怀疑是否有可能分析灭绝的古人类的远古DNA。manbet手机版通过广泛的技术发展,Svante Pääbo在这一具有挑战性的领域设立了新的严格标准,并成功地获得了我们已灭绝的近亲尼安德特人的基因组序列。manbet手机版随后,他又通过从一个小指骨标本中提取的基因组数据,轰动地发现了另一种已灭绝的古人类——丹尼索瓦人。manbet手机版Svante Pääbo的研究进一步证实,智人在共存时期与尼安德特人和丹尼索瓦人混合,导致古代人的DNA渗入现代人类。manbet手机版影响当今人类生理机能的古老基因变异的显著例子已经在一个高度动态的研究领域中得到证实。manbet手机版通过他突破性的发现,Pääbo打开了一扇了解人类进化历史的新窗口,揭示了古代古人类进化和混合的意想不到的复杂性,同时也为更好地理解使我们成为独一无二的人类的基因特征提供了基础。
manbet手机版我们的起源问题长期以来一直引起人类的兴趣,一直是人们强烈兴趣和讨论的焦点。manbet手机版目前所有生物体的进化都始于数十亿年前,第一批灵长类动物出现在5500 - 6500万年前。manbet手机版据估计,旧大陆猴子和类人猿的最后一个共同祖先生活在大约2500万年前,人类和黑猩猩的最后一个共同祖先生活在大约600万年前。manbet手机版属manbet手机版人类manbet手机版大约在三百万年前进化而来manbet手机版直立人manbet手机版进入180万年前的场景。manbet手机版和大多数进化过程一样,我们的早期历史以众多的基因分支为特征,其中许多最终都不成功。manbet手机版然而,manbet手机版直立人manbet手机版比其他古人类物种存活的时间更长,是已知的第一个离开非洲的古人类物种,随后的进化导致了manbet手机版人类neandertalensismanbet手机版即我们的远古亲戚尼安德特人。
manbet手机版在很长一段时间里,人类进化的研究依赖于对古代骨骼遗迹的分析、它们的形态特征以及对工具和其他考古器物的检查。manbet手机版第一个被确认为尼安德特人的骨骼遗骸于1856年在德国的尼安德特山谷被发现。manbet手机版从那时起,来自多个尼安德特人的标本在欧亚大陆的广阔地理范围内被发现。manbet手机版尼安德特人大约在40万到3万年前居住在欧洲和亚洲,之后他们灭绝了。manbet手机版在解剖学上现代人类,manbet手机版智人manbet手机版约30万年前首次出现在非洲。manbet手机版大约6万到7万年前,从非洲到中东的移民导致了manbet手机版智人manbet手机版横跨欧亚大陆。manbet手机版因此,manbet手机版智人manbet手机版与尼安德特人在欧亚大陆共存了至少2万年,可能更长。manbet手机版然而,它们相互作用的性质是一个长期争论的问题。manbet手机版越来越清楚的是,需要通过基因分析来阐明我们和我们的尼安德特亲戚之间的关系。
manbet手机版使用基因工具来研究当代人类是如何相互关联的,并阐明我们的进化起源的第一步是由艾伦·威尔逊迈出的。manbet手机版在一项关键研究中,他对当前非洲人和非非洲人群体的线粒体DNA (mtDNA)进行了限制性内切酶分析,这是人类DNA中最容易获取的部分,因为它在每个细胞中都有很高的拷贝数。manbet手机版他的研究结果表明,所有亚种都有一个共同的非洲起源manbet手机版智人manbet手机版.manbet手机版这些结果为人类进化的“走出非洲”理论奠定了基础。manbet手机版然而,由于mtDNA只在母系侧遗传,而且它仅由16500个碱基对(bp)组成,而核基因组有30亿bp,因此这些结果并不确定。manbet手机版因此,需要更全面的数据,包括来自人类核基因组的数据。
manbet手机版人类核基因组研究的跳板是人类基因组计划,该计划于1990年由一个大型国际财团发起。manbet手机版目的是测序和注释一个完整的单倍体核基因组,并为未来的研究生成一个人类参考地图。manbet手机版2001年,发表了两篇描述人类核基因组序列的里程碑式论文[2,3],其余复杂区域直到今年[4]才完成。manbet手机版参考基因组可从多个来源向公众提供,包括加州大学圣克鲁兹分校、国家生物技术信息中心、GenBank和ensemble。manbet手机版随后,来自不同人群的更多人类基因组被报道,包括来自1000基因组计划联盟[5]和Simons基因组多样性计划[6]的基因组,为理解人类群体遗传学和探索我们的进化历史提供了重要资源。manbet手机版然而,要弄清我们与已灭绝的古人类(如尼安德特人)之间的关系,需要的不仅仅是对当代人类DNA的推断研究。manbet手机版这需要从灭绝物种中提取非常古老的DNA并进行测序,这是一项似乎无法实现的艰巨任务。
manbet手机版Svante Pääbo在乌普萨拉大学学习医学的时候,从很小的时候就对埃及学产生了浓厚的兴趣。manbet手机版在Per a . Peterson的小组进行腺病毒及其与免疫系统相互作用的研究生研究期间,他秘密从事了一个业余项目,旨在从木乃伊标本中分离DNA。manbet手机版他成功地从一具2400年前的木乃伊样本中克隆了一个DNA库,并用人类重复序列对其进行筛选,在克隆中发现了人类DNA。manbet手机版然而,Pääbo很快意识到,研究古代DNA存在技术挑战,他后来承认,第一篇论文中描述的结果很可能受到了来自当代人类DNA的污染。manbet手机版因此,他专注于改进技术,这最好是通过分析来自非人类物种的古代DNA,污染更容易被检测到。manbet手机版1987年,在Zürich大学沃尔特·沙夫纳的实验室做了一段短暂的研究员后,Pääbo转到加州大学伯克利分校,在艾伦·威尔逊的实验室做博士后,研究古代DNA。
manbet手机版古代DNA分析的方法论挑战
manbet手机版在威尔逊实验室,以及在他职业生涯的未来几十年里,Svante Pääbo不知疲倦地致力于开发从古代标本中提取、测序和分析DNA的协议。manbet手机版尽管这项工作的大部分是在其他物种上进行的,但他的最终目标是从尼安德特人的骨头中提取的DNA中获取序列信息。manbet手机版在20世纪80年代中期,由Kary Mullis发明并于1993年获得诺贝尔化学奖的聚合酶链式反应(PCR)彻底改变了该领域,极大地促进了微量DNA的分析。狗万世界杯manbet手机版然而,恢复受污染DNA的风险仍然存在,甚至增加了。manbet手机版1988年,Pääbo和Wilson率先将PCR应用于古代遗迹[8]。manbet手机版Pääbo敏锐地意识到与非常古老的骨骼样本的DNA分析相关的技术挑战。manbet手机版他和其他人已经证明,来自晚更新世时期的骨骼受到了微生物和现代人处理标本的DNA的严重污染,而内源性DNA,即使存在,也只发现了微量。manbet手机版该领域的几个小组都很活跃,但许多研究都受到了污染,导致了虚假的结果。manbet手机版Svante Pääbo果断地解决了这一问题,设计了专门的洁净室,最大限度地减少污染,并强调需要由其他实验室独立复制结果。
manbet手机版测定样本中古DNA含量的方法也是迫切需要的,同时也需要更好地了解古DNA的生化特性。manbet手机版在古代样本中观察到的DNA损伤类型包括核苷酸的破碎和修饰,这是由氧化过程引起的,如胞嘧啶残基脱氨生成尿嘧啶。manbet手机版几十年来,Pääbo的小组描述了与分析古代DNA相关的挑战,并提供了解决方案[9-11]。manbet手机版该小组还开发了使用硅基纯化方法[12]从古代样本中提取DNA的方法,并证明了某些氨基酸的外消旋化程度可以用来确定样本是否含有内源性DNA[13]。
manbet手机版理论衰减率的研究对于了解旧DNA的命运也很重要。manbet手机版Pääbo曾与托马斯·林达尔(Thomas Lindahl)在伦敦帝国癌症研究基金(Imperial Cancer Research Fund)短暂合作过一段时间,他在一篇文章中认为,可以成功分析数万年甚至数十万年的样本,而更老的样本预计不会产生结果[14]。manbet手机版然而,DNA储存的条件对其衰减率有很大的影响,低温的破坏较小。manbet手机版例如,最近对从永久冻土中发现的一颗100万年前猛犸象牙齿的DNA测序说明了这一点[15]。
manbet手机版这些发现
manbet手机版最初的突破是:尼安德特人线粒体DNA
manbet手机版尽管有了聚合酶链反应(PCR)的发明,但在古骨中发现的少量残留DNA是否能够抵抗化学修饰和碎片化数万年,足以抵御来自微生物生长的更近的DNA和当今人类DNA污染的竞争,仍然是一个非常值得怀疑的问题。manbet手机版然而,Svante Pääbo在1990年被慕尼黑大学录取,他决心实现这一目标。manbet手机版最初的尼安德特人骨骼遗骸是1856年在Düsseldorf附近的尼安德特山谷的费尔德霍弗洞穴中发现的。manbet手机版这个“模式标本”的残骸保存在德国波恩的莱茵博物馆。manbet手机版当Pääbo开始他雄心勃勃的探索研究尼安德特人的基因组时,这是他第一次寻找珍贵遗骸的地方,他需要这些遗骸作为材料的来源。manbet手机版幸运的是,他很受欢迎,在合作的基础上,他得到了一块尼安德特人的肱骨。
manbet手机版线粒体DNA比核DNA有更大的机会在年老的骨骼中保存下来,因为每个细胞包含数百到数千个线粒体DNA副本,但每个核DNA片段只有两个副本(一个是母系的,一个是父系的)。manbet手机版因此,Pääbo选择mtDNA作为他研究的第一个尼安德特人序列目标。manbet手机版两个引物定义了人类mtDNA控制区高变量部分105 bp的片段用于PCR扩增,并对两次独立实验得到的片段进行克隆和测序。manbet手机版他获得的初始mtDNA序列有61个核苷酸长。manbet手机版大多数克隆体都含有在现代人类中未见过的变异。manbet手机版采取了广泛的预防措施,并进行了对照实验,以确保序列是内源性的。manbet手机版用一块新的骨头重复了这些实验,得到了相同的结果。manbet手机版一块尼安德特人的骨头也被送到宾夕法尼亚州立大学人类学遗传学实验室的马克·斯托金(Mark Stoneking)那里,在一个独立的实验室里进行复制。manbet手机版然后对重叠的DNA片段进行扩增、克隆和测序,以重建一个更长的序列。manbet手机版共有123个克隆,代表13个片段,每个克隆至少测序两次,将尼安德特人mtDNA序列扩展到379个核苷酸[16]。
manbet手机版为了研究现代人和尼安德特人之间的关系,研究人员将尼安德特人mtDNA的379个核苷酸与2051个人类和59个黑猩猩mtDNA序列进行了比较。manbet手机版该序列显示在欧洲人、非洲人、亚洲人、美洲土著人和澳大利亚/大洋洲人的变异之外manbet手机版智人manbet手机版(manbet手机版图1manbet手机版).
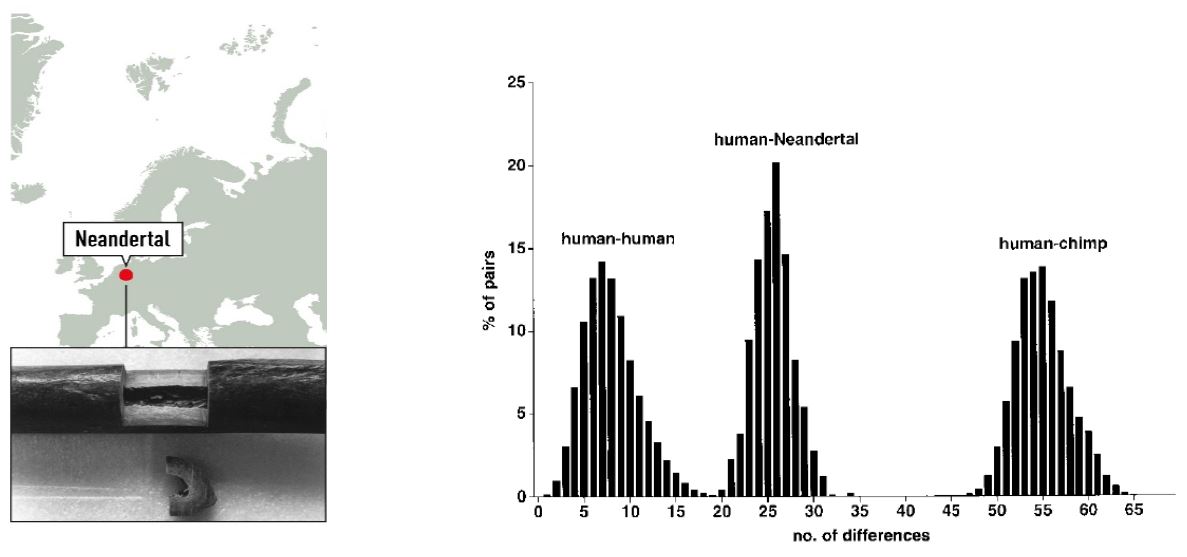
manbet手机版图1所示。manbet手机版左:尼安德特人遗址的地图和模式标本的右肱骨样本。manbet手机版右图:人类、尼安德特人和黑猩猩之间两两序列差异的分布(Krings et al. 1997)。
manbet手机版系统发育树模型表明,现在的mtDNA基因库起源于非洲,这与威尔逊之前的发现一致,携带这种mtDNA的共同祖先生活在12万至15万年前。manbet手机版根据mtDNA分析,从尼安德特人和解剖学上的现代人共同祖先存在的时间估计是现在的四倍;manbet手机版55万到69万年。manbet手机版结果表明,尼安德特人在没有向现代人类提供mtDNA的情况下就灭绝了。
manbet手机版由于mtDNA只通过母系遗传,只能由雌性遗传给下一代,它只能给出我们进化史的有限图景。manbet手机版由于mtDNA反映的是女性的一面,所以只有当女性在两组之间移动时,mtDNA才会显示出来。manbet手机版由于其严格的母系遗传和缺乏重组,如果携带者女性只有儿子,mtDNA变异也可能偶然消失。manbet手机版因此,mtDNA分析的信息量不如核基因组的研究。manbet手机版对尼安德特人mtDNA的测序和分析的结果并不排除尼安德特人为现代人类贡献了其他基因的可能性。manbet手机版对尼安德特人mtDNA的测序证明,我们有可能对我们最近灭绝的近亲4万年前的骨头进行DNA测序。manbet手机版然而,需要更多的骨骼来巩固和扩展这些令人兴奋的结果。
manbet手机版克罗地亚的文迪加因发现尼安德特人的重要骨骼而闻名。manbet手机版就像德国的尼安德特人遗址一样,它含有石灰石洞穴,由于其轻微的基础化学环境,特别适合保存DNA。manbet手机版这些标本保存在萨格勒布的克罗地亚科学和艺术学院古生物学和地质研究所。manbet手机版Pääbo访问了该研究所,得到了15个尼安德特人骨头的小样本。manbet手机版氨基酸分析表明,7个标本可能含有足够数量的尼安德特人DNA。manbet手机版最有希望的样本是碳-14年代测定法,显示有42000年的历史。
manbet手机版2000年,一个英国小组公布了第二个尼安德特人mtDNA序列,来自北高加索的Mezmaiskaya洞穴[17]。manbet手机版此后不久,Pääbo的研究小组发表了Vindija标本的第三个mtDNA序列,从而对尼安德特人[18]的遗传多样性进行了估计。manbet手机版艾伦·威尔逊曾指出,黑猩猩、大猩猩和红毛猩猩的基因变异程度比现代人更大,尽管它们的数量要少得多,这表明了这一点manbet手机版智人manbet手机版从一个较小的人口扩展而来。manbet手机版对三个尼安德特人mtDNA序列的分析得出了类似的结果,表明这些人来自一个随后经历了扩张的小群体。
manbet手机版2004年,Pääbo扩大了尼安德特人mtDNA变异的研究范围,增加了来自欧洲不同地理位置的另外四个尼安德特人样本和五个早期的尼安德特人样本manbet手机版智人manbet手机版[19]。manbet手机版基于对mtDNA的分析,研究小组得出结论,尼安德特人对早期现代人的基因贡献很大的可能性不大,尽管他们不能排除较小的基因贡献的可能性。
manbet手机版第一个一百万个尼安德特人核苷酸
manbet手机版Pääbo,他现在已经在德国莱比锡建立了马克斯·普朗克进化人类学研究所,在新测序技术的出现刺激下,他转向了核基因组。manbet手机版使用454生命科学公司(454 Life Sciences)提供的一项当时的新技术,创建了单链DNA文库,侧边是普通的连接体,通过珠基乳剂PCR扩增单个分子,然后进行邻苯二氢测序[20]。manbet手机版与之前使用的PCR、克隆和测序方法相比,该测序平台具有几个优点。manbet手机版重要的是,它每次运行产生数十万个读取,因此提供了高吞吐量的格式。manbet手机版它还可以直接对样本中的所有DNA进行测序,而无需通过PCR选择序列,并避免了细菌克隆的需要,因为在细菌克隆中会丢失大量的模板分子。manbet手机版它进一步排除了PCR中的模板竞争,因为每个分子都是单独扩增的。manbet手机版它的读取长度为100-200个核苷酸,与保存在古骨中的DNA的平均长度相吻合。manbet手机版它还允许推断不同核苷酸错配的频率,因为每个测序产物都来自一个已知方向的原始单链模板分子。
manbet手机版Pääbo的团队测试了来自欧洲和西亚不同地点的70多个尼安德特人的骨头和牙齿样本,以估计生物分子保存情况。manbet手机版大多数样本不太可能含有可回收的内源性DNA,但一些样本含有高水平的氨基酸和低水平的氨基酸外消旋化,这些因素表明检索古DNA的机会是合理的。manbet手机版通过PCR和mtDNA测序来评估现代人类DNA的污染水平。manbet手机版结果表明,在不同的样本之间,差异很大。manbet手机版克罗地亚Vindija洞穴的一块骨头突出了90%的mtDNA片段来自尼安德特人。
manbet手机版这块骨头被选作高通量测序。manbet手机版使用BLASTN程序,共恢复了254,933个独特序列,并将其与人类、黑猩猩和老鼠的基因组序列和GenBank数据库进行了比对。manbet手机版研究人员分析了与人类或黑猩猩核基因组最接近的至少30个碱基对的序列。manbet手机版总的来说,0.04%的尼安德特人核基因组序列被reads覆盖。
manbet手机版通过高通量测序产生的第一个一百万个核苷酸于2006年发表[21]。manbet手机版爱德华·鲁宾(Edward Rubin)的团队与Pääbo合作,尝试了一种基于克隆同一块骨头的替代策略,产生了65000个核苷酸[22]。manbet手机版总之,这些结果表明,DNA序列确实可以从尼安德特人的核基因组中生成,为进一步的研究提供了概念证明。manbet手机版随后的分析表明,高通量测序生成的数据集受到了现代人类DNA[23]的污染,这推动了后续工作中进一步的方法改进。manbet手机版Pääbo在他的文章[21]的最后一段中指出,随着技术的进一步改进,可以设想实现尼安德特人完整基因组(30亿碱基对)的一倍覆盖率。manbet手机版Pääbo最后宣布,他已经启动了一个项目,旨在在两年内获得尼安德特人基因组的初步草案。
manbet手机版尼安德特人基因组序列草图
manbet手机版为了实现他的大胆计划,Pääbo需要更多的骨头。manbet手机版从Vindija收集的其他标本是与柏林-勃兰登堡科学院和克罗地亚科学和艺术学院合作获得的。manbet手机版其中的三个标本,每个大约有4万年前,包含超过1%的尼安德特人DNA,并被选为尼安德特人核基因组测序。manbet手机版其他来源还包括尼安德特山谷、高加索地区的梅兹迈斯卡亚洞穴和西班牙的El Sidrón洞穴。manbet手机版在洁净室条件下构建并添加了特定的适配器,其中包含一个尼安德特人特定的序列密钥,可以明确地识别每个序列,确定它们来自尼安德特人[9]骨的提取物,并进行了额外的关键方法改进。manbet手机版文库制备方案得到改进,可读DNA[24]的产量提高了数百倍,通过使用优先切割文库中的细菌序列的限制性内切酶,提高了尼安德特人与微生物DNA的比例。
manbet手机版2008年,该团队开始使用新兴的测序技术,可以实现更高的吞吐量。manbet手机版他们获得了10亿个长度为30-70个核苷酸的序列。manbet手机版测序平台的改变具有挑战性,因为需要开发新的算法来分析和映射数据[25]。manbet手机版在分析古DNA时,一种比对方法被实现,该方法考虑了核苷酸错配的类型(C到T的转变,特别是在读取结束时)。manbet手机版参考人类和黑猩猩的基因组以及推断出的人类和黑猩猩共同祖先的基因组序列被用于比较分析。
manbet手机版现在需要不同的专门知识来分析数据,包括人口遗传学家,因此成立了一个约50名科学家组成的联盟。manbet手机版其中,大卫·赖希、尼克·帕特森、蒙哥马利·斯拉特金和吉姆·穆利金都是关键贡献者,还有后来被邀请加入团队的拉斯穆斯·尼尔森和翟薇薇。manbet手机版从这三个个体的DNA中得到的序列信息包含了超过40亿个核苷酸,这些核苷酸随后被绘制出来并进行分析。manbet手机版2008年,使用8341个mtDNA序列重建了完整的16,565个核苷酸mtDNA。manbet手机版对组装序列的分析明确地确定尼安德特人mtDNA不在现存人类mtDNA的变异范围内,并允许估计两个mtDNA谱系之间的差异为66万年[26]。
manbet手机版2010年,在一份突破性的出版物中,Pääbo实现了无法实现的目标,并报道了尼安德特人核基因组序列[27]的草稿。manbet手机版他大胆的计划实现了。manbet手机版5今天manbet手机版智人manbet手机版来自世界不同地区的基因组也进行了测序,以帮助进行比较数据分析。manbet手机版使用三种方法来估计数据中现代人类DNA的污染。manbet手机版首先,我们使用了尼安德特人和现代人类mtDNA之间已知的不同位置。manbet手机版其次,对于来自女性尼安德特人的骨头,现代人类男性DNA污染是通过寻找Y染色体非重组部分的独特片段来估计的。manbet手机版最后,研究人员使用了现代人与黑猩猩和尼安德特人不同的核基因组位点。manbet手机版这三种方法得出的污染估计都不到1%。
manbet手机版从另外三个覆盖晚期尼安德特人大部分地理范围的遗址中挖掘出的尼安德特人的骨头中产生了少量的DNA序列:西班牙阿斯图里亚斯的El Sidròn,德国尼安德特谷的Feldhofer洞穴和俄罗斯高加索地区的Mezmaiskaya洞穴。manbet手机版估计每个标本的DNA差异与人类参考基因组显示,它们与Vindija个体没有显著差异。manbet手机版因此,来自欧亚大陆西部大部分地区的尼安德特人似乎与现代人类有着同样的亲缘关系。manbet手机版尼安德特人和现代人类核DNA序列的平均差异估计为825,000年[27]。
manbet手机版关于尼安德特人和解剖学上的现代人之间是否发生过杂交的问题,现在可以通过核基因组序列分析直接进行调查了。manbet手机版当Pääbo调查尼安德特人与当今人类的亲缘关系时,他最初使用了两个欧洲裔美国人,两个东亚人和四个西非人,令人惊讶的是,他发现尼安德特人与欧洲人和东亚人的亲缘关系是一样的,与非非洲人的亲缘关系要比非洲人的亲缘关系密切得多。manbet手机版将法国人、汉族、巴布亚人、约鲁巴人和桑人的基因组序列相加进行扩展分析,得出了相同的结论。manbet手机版尼安德特人与非非洲人的关系比与非洲人的关系更密切,这一发现最容易用尼安德特人与非非洲人的祖先在共同生活期间的基因流动(渗透)来解释。manbet手机版作为基因渗入的一个独立指标,许多在非洲以外比在非洲内部变异更大的基因组区域被证明代表尼安德特人序列。
manbet手机版尼安德特人与中国人和巴布亚人的关系就像与法国人的关系一样密切,尽管在形态上可辨认的尼安德特人只存在于欧洲和西亚的记录中。manbet手机版这可以用早期的混合来解释manbet手机版智人manbet手机版他们的祖先是现在的非非洲人尼安德特人,可能在他们扩张到欧亚大陆之前生活在中东。manbet手机版数据显示,在欧亚大陆的人类基因组中,有1%到4%来自尼安德特人。
manbet手机版进一步的分析发现,在所有测序的现代人类中都存在变异,而在尼安德特人中却没有。manbet手机版共有78个核苷酸取代改变了基因的蛋白质编码能力,被发现是固定的派生状态manbet手机版智人manbet手机版而尼安德特人则继承了祖先的地位。manbet手机版因此,在过去几十万年的人类进化中,相对较少的氨基酸变化被固定下来。manbet手机版研究人员还发现,与尼安德特人相比,现代人的一些潜在的非编码调控替代是固定的。manbet手机版随着基因组序列数据的积累,人类独特特征的目录不断扩大,形成了一个非常活跃的研究领域的基础。
manbet手机版一种新的古人类的发现:丹尼索瓦人
manbet手机版通过对尼安德特人的基因组进行测序,Pääbo证明了一种已灭绝的晚更新世古人类的基因组序列可以可靠地恢复和分析,开创了研究我们进化史的新方法。manbet手机版自从我们最亲近的进化亲戚灭绝以来,DNA需要特殊的环境才能存留很长一段时间。manbet手机版降解程度随着温度和土壤条件(如酸性)的增加而增加,大多数早期古人类的骨骼来自非洲和欧亚大陆的赤道和热带地区,那里维持DNA完整性的条件很差。
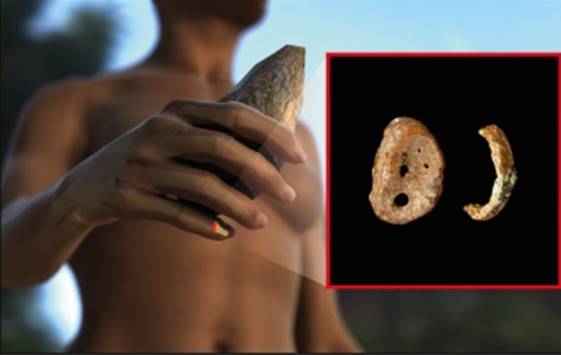
manbet手机版图2。manbet手机版手的远端phalynx,在Denisova洞穴中发现。
manbet手机版然而,考古证据表明,古人类也生活在高纬度地区,那里的DNA保存潜力更大。manbet手机版其中一个这样的地区是西伯利亚南部的阿尔泰山脉,古人类可能在12.5万多年前就在那里居住过。manbet手机版在阿尔泰的大多数遗址中都不存在适合进行形态分类的骨骼标本,但已经发现了一些人类骨骼的小碎片,如牙齿和骨头碎片。
manbet手机版2008年,在俄罗斯阿尔泰山脉的丹尼索瓦洞穴(Denisova Cave)中挖掘出了一个幼年古人类第五指的远端指骨,其地层可以追溯到4.8万至3万年前。manbet手机版Pääbo的研究小组从这块指骨上制成粉末并提取DNA,发现DNA保存得非常完好。manbet手机版引物扩展捕获最初用于分离mtDNA片段。manbet手机版这些被测序和组装成一个完整的mtDNA序列,平均覆盖156倍[28]。manbet手机版几种不同的方法被用来评估mtDNA序列的可靠性。manbet手机版例如,第二个DNA提取物被用于霰弹枪测序,结果组装的mtDNA序列与引物扩展捕获获得的第一个序列相同。
manbet手机版该序列与54个现代人类mtDNA、一个来自早期现代人的更新世晚期mtDNA、6个完整的尼安德特人mtDNA、一个倭黑猩猩mtDNA和一个黑猩猩mtDNA相匹配。manbet手机版然而尼安德特人的mtDNA不同于manbet手机版智人manbet手机版在平均202个核苷酸位置上,手指骨样本的平均差异为385个位置,而黑猩猩的平均差异为1462个位置。manbet手机版因此,与尼安德特人相比,来自未知古人类的mtDNA与现代人的差异要大得多。manbet手机版2010年,对这些序列的系统发育分析证实了一个惊人的事实:这是一种独特的人类inin[28],被命名为Denisova。manbet手机版丹尼索瓦古人类、尼安德特人和现代人共同拥有的最新mtDNA祖先的年代估计约为100万年前,是现代人和尼安德特人最新共同mtDNA祖先的两倍。
manbet手机版这一发现的意义是巨大的。manbet手机版Pääbo发现了一种全新的古人类,与尼安德特人和manbet手机版智人manbet手机版.manbet手机版此外,这一发现完全是通过古代DNA的检索和测序获得的,没有任何可用的形态学信息。manbet手机版Pääbo的团队继续按照严格的程序,从丹尼索瓦人指骨中提取DNA,对核基因组进行测序。manbet手机版他们将这些序列映射到人类和黑猩猩的参考基因组,以及这些物种的推断祖先基因组,生成了丹尼索瓦人的基因组序列,覆盖率约为[29]的1.9倍。
manbet手机版研究了丹尼索瓦人的基因组与尼安德特人和现代人的关系。manbet手机版尽管该团队已经表明,丹尼索瓦人的mtDNA与现代人mtDNA的差异大约是尼安德特人mtDNA差异的两倍,但丹尼索瓦人与现代人核基因组的平均差异与尼安德特人的相似但不同,这表明他们是姐妹群体。manbet手机版据估计,尼安德特人和丹尼索瓦人的DNA序列平均在64万年前就有差异,而与今天非洲人的DNA序列则在约80.4万年前就有差异。
manbet手机版研究人员对四组基因组进行了比对,一组是非洲基因组,一组是欧亚基因组(法国基因组),一组是古人类基因组(尼安德特人或丹尼索瓦人)和黑猩猩基因组。manbet手机版尼安德特人和丹尼索瓦人的基因组与法国人基因组的匹配程度都高于非洲人的基因组,但欧亚人基因库的古老部分与丹尼索瓦人的关系似乎不如与尼安德特人的密切。manbet手机版与非洲个体相比,欧亚个体与丹尼索瓦人表现出更大的相似性,这一事实表明,在基因从尼安德特人流入非非洲人之前,丹尼索瓦人与尼安德特人有一些共同的历史。
manbet手机版因此,丹尼索瓦人似乎没有直接参与假定的基因从尼安德特人流入欧亚人的过程。manbet手机版然而,当丹尼索瓦人的基因组与来自53个群体的938个现代人的基因组的关系进行研究时,发现巴布亚人和布干维尔岛民与几乎所有个体都不同。这些人的基因分型在642,690个单核苷酸多态性位点上。
manbet手机版通过分析更多的基因组,进一步探索了这一点,支持了这样的结论:丹尼索瓦人的遗传物质对今天美拉尼西亚人的基因组贡献了约4-6%。manbet手机版这令人惊讶,因为这表明丹尼索瓦人的地理范围很广,横跨欧亚大陆的东部和南部。manbet手机版根据这些结果,Pääbo和他的团队提出,丹尼索瓦人是尼安德特人的姐妹群体,其人口分化时间是尼安德特人和人类共同祖先的一半到三分之二。manbet手机版丹尼索瓦人和尼安德特人分化后,尼安德特人的基因流到了今天非非洲人的祖先身上。manbet手机版后来丹尼索瓦人和美拉尼西亚人的祖先混种,这并没有影响到其他非非洲人。manbet手机版图3manbet手机版).
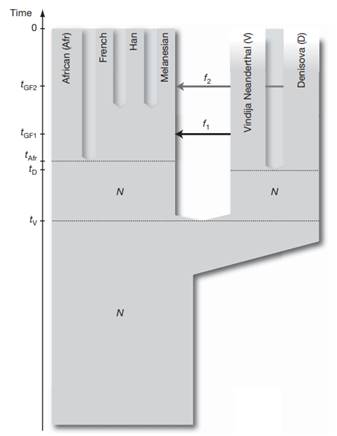
manbet手机版图3manbet手机版.manbet手机版与数据相吻合的种群历史模型。manbet手机版N表示有效种群大小,t表示种群分离时间,f表示基因流动量,tGF表示基因流动量时间(Reich et al. 2010)。
manbet手机版2012年,Pääbo从2008年发现的原始指骨中获得了高覆盖率(>30倍)的丹尼索瓦人基因组序列,使用旨在增加[30]测序DNA分子数量的单链文库。manbet手机版2010年,在丹尼索瓦洞穴(Denisova Cave)发现了另一块古人类骨骼,这次是近端脚趾指骨。manbet手机版2014年,Pääbo的团队从这块骨头中生成了完整的、高质量的基因组序列,表明它来自阿尔泰山脉的尼安德特人[31]。manbet手机版这证实了尼安德特人的活动范围一直延伸到西伯利亚。manbet手机版他们将这些基因组与现有覆盖率较低的尼安德特人基因组进行了比较,包括来自高加索地区梅兹迈斯卡亚洞穴的一个基因组,以及25个高覆盖率的当代人类基因组。manbet手机版根据这些数据,>Pääbo的小组估计,尼安德特人和丹尼索瓦人的人口分裂时间为38万至47万年前,现代人和尼安德特人/丹尼索瓦人的分裂时间为55万至76万年前。manbet手机版此外,他们在尼安德特人的血统中发现了近亲繁殖的证据,而且多样性水平很低,这表明尼安德特人的种群规模很小。manbet手机版他们将目前欧亚大陆基因组中尼安德特人祖先的估计提高到1.5-2.1%,并证明了尼安德特人和欧洲人之间存在更复杂的混合历史manbet手机版智人manbet手机版.manbet手机版研究小组还发现了从阿尔泰地区的尼安德特人到丹尼索瓦人的基因流动的证据。
manbet手机版古人类和古人类的爆发manbet手机版智人manbet手机版基因组se-quences
manbet手机版随着他开创性的发现,Pääbo建立了一门全新的科学学科,现在被称为古基因组学。manbet手机版大量的基因组序列数据已经从我们已经灭绝的古人类亲属,以及古代manbet手机版智人manbet手机版这个领域正在迅速发展。manbet手机版2014年,Pääbo的团队使用杂交方法富集了来自西班牙和克罗地亚的两个尼安德特人基因组的蛋白质编码部分的DNA,并与西伯利亚尼安德特人基因组一起分析了序列。manbet手机版尼安德特人的基因多样性被证明比今天的人类更低和不同,这表明尼安德特人的人口数量小,彼此之间是孤立的。manbet手机版进一步表明,在尼安德特人进化谱系中,涉及骨骼形态的基因变化比预期的要大,而在现代人类谱系[32]中,涉及色素沉着和大脑功能的基因变化更大。
manbet手机版2017年,Pääbo的团队对第二个尼安德特人基因组进行了高覆盖率测序(>30倍),该基因组来自克罗地亚Vindija洞穴的一个尼安德特人,距今约5万年前[33]。manbet手机版她的两个基因组拷贝之间每10000个碱基对携带1.6个差异,比现在的人类要少,这支持了尼安德特人的人口规模小的观点。manbet手机版2018年,Pääbo和珍妮特·克尔索应用了一种使用次氯酸盐处理的方法,从生活在3.9 - 4.7万年前的5名尼安德特人(被称为晚期尼安德特人[34])中生成了低覆盖率(1-2.7倍)的基因组序列。manbet手机版这使得尼安德特人基因组序列的数量翻了一番,并使尼安德特人的群体基因组研究成为可能。manbet手机版该小组观察到晚期的尼安德特人彼此相似,但与高加索地区的较老的尼安德特人不同,这表明较老的尼安德特人谱系被新的尼安德特人群体所取代。manbet手机版这种人口更替可能发生在尼安德特人人口历史的末期。
manbet手机版2015年,Pääbo的小组对来自丹尼索瓦洞穴[35]的另外两个丹尼索瓦个体的mtDNA基因组和核基因组进行了特征分析。manbet手机版他们发现,丹尼索瓦人的细胞核基因组与原始的丹尼索瓦人基因组形成了一个分支,而其中一个新测序的丹尼索瓦人的mtDNA基因组比其他的丹尼索瓦人更分散,他们解释说,这表明丹尼索瓦人在该地区生活了很长一段时间。manbet手机版2017年,他们对第四个丹尼索瓦人的mtDNA和47 Mb基因组序列进行了测序,得出的结论是,这个人比其他两个丹尼索瓦人要老得多,加强了丹尼索瓦人很可能在丹尼索瓦洞穴附近生活了很长时间的观点[36]。manbet手机版随后,从丹尼索瓦洞穴[37]的另一块骨头碎片中分离出的低覆盖率(2.6倍)基因组中,又有了一个显著的发现。manbet手机版根据整个基因组的杂合度水平,研究小组得出结论,这个人是尼安德特人母亲和丹尼索瓦人父亲的第一代后代。
manbet手机版Pääbo的团队随后发表了第三个高覆盖率的尼安德特人基因组[38],来自阿尔泰山脉Chagyrskaya洞穴的一个个体。manbet手机版她与欧亚大陆西部的尼安德特人的亲缘关系比与更早生活在丹尼索瓦洞穴的尼安德特人的亲缘关系更大,这与更古老的尼安德特人谱系的灭绝和后来欧亚大陆西部尼安德特人向东传播的可能性一致。manbet手机版基于该基因组中众多的纯合子区域,他们推断西伯利亚尼安德特人生活在相对孤立的少于60个个体的群体中。manbet手机版相反,来自欧洲的尼安德特人,来自阿尔泰山脉的丹尼索瓦人,以及古代现代人似乎生活在更大规模的人口中。
manbet手机版在一项特别了不起的技术成就中,Pääbo的团队从两个样本中恢复了核DNA序列,这些样本来自28个古人类个体,发现于西班牙阿塔普埃卡山脉的西马德洛斯胡索斯,可追溯到大约43万年前。manbet手机版这些人与尼安德特人有一些共同的形态特征,但他们与尼安德特人和丹尼索瓦人的关系尚不清楚。manbet手机版Pääbo的研究小组表明,西马德洛斯胡索古人类与尼安德特人有关,而不是丹尼索瓦人。
manbet手机版除了Pääbo对已经灭绝的古人类基因组的突破性发现外,对古代DNA降解的深入理解、高通量DNA测序技术和强大的计算资源的结合,导致了对古代现代人类基因组的检索。manbet手机版一个早期的例子是一个4000岁的爱斯基摩人的基因组序列,2010年由Eske Willerslev[40]小组报道。manbet手机版为了追踪已经灭绝的古人类在早期现代人中渗透的证据,Pääbo的团队从来自西西伯利亚的45000年前的现代人[41]和来自罗马尼亚的40000年前的现代人[42]中生成了基因组序列。manbet手机版两者都显示了最近与尼安德特人基因组传入的证据。
manbet手机版现在,从不同地理位置和不同时期发现的古人类标本中获得了丰富的基因组数据资源,而且数据正在迅速积累。manbet手机版Svante Pääbo从我们已灭绝的近亲中提取的古老基因组序列被广泛用于比较分析,以阐明种群扩散和人类适应历史。manbet手机版古人类素样本的稀缺限制了基因组序列的可用性。manbet手机版最近,Svante Pääbo, Matthias Meyer及其同事发现,可以从更新世沉积物中提取古人类的mtDNA[43,44]以及核DNA[45],为研究灭绝古人类的时间地理分布和种群动态提供了全新的可能性。manbet手机版Pääbo发现的高覆盖率的古人类基因组序列构成了必要的支架,沉积物和骨骼中不太完整的全基因组序列数据可以在此基础上进行绘制。manbet手机版高覆盖率的基因组也为与当今人类进行令人兴奋和重要的比较分析奠定了基础。
manbet手机版影响
manbet手机版深入了解我们古老的进化史
manbet手机版Pääbo的发现对理解人类进化史产生了深远的影响,也刺激了该领域的研究。manbet手机版我们现在知道,至少有两个不同的古人类群体,尼安德特人和丹尼索瓦人居住在欧亚大陆,当时解剖学上的现代人(manbet手机版智人manbet手机版)起源于非洲。manbet手机版Pääbo的团队在丹尼索瓦洞穴中发现了三颗牙齿,它们携带的基因组序列与原始丹尼索瓦人基因组提取的指骨的基因组序列高度相似,但却截然不同[29,35,36]。manbet手机版因此,牙齿和指骨很可能来自同一古人类种群的不同个体。manbet手机版形态特征与尼安德特人和现代人的牙齿不同,进一步表明丹尼索瓦人的进化史与尼安德特人和现代人截然不同。
manbet手机版从不同的地理位置和时间点检索到的与我们最接近的已灭绝近亲的基因组序列的可用性,为它们的分布、种群结构和混合提供了线索。manbet手机版Pääbo的发现被用来改进“走出非洲”模型,在这个模型中,所有现代人类都以一条不间断的线追溯到非洲。manbet手机版他的数据显示,古代现代人、尼安德特人和丹尼索瓦人之间发生了杂交,导致少量的古代基因组片段保留在现代人身上。manbet手机版非非洲人基因组包含约1-2%的尼安德特人遗传物质。manbet手机版渗入的古DNA片段在个体之间的位置是不同的,总共约40%的尼安德特人基因组已经在现代人身上发现。manbet手机版有趣的是,一些基因组区域完全没有来自尼安德特人或丹尼索瓦人祖先的信号,被称为“古沙漠”,这表明在这些区域的基因渗透是不可容忍的[46-48]。manbet手机版除了尼安德特人的基因序列,美拉尼西亚人从古丹尼索瓦人[49]获得了4-6%的遗传物质。manbet手机版从尼安德特人和丹尼索瓦人到非非洲人的多波渗透已经被指出,反映了古代现代人和灭绝的古代血统之间的许多杂交繁殖事件[48,50]。manbet手机版此外,丹尼索瓦人遗产的比例在太平洋族群[51]之间差异显著。
manbet手机版DNA在古骨中存活需要特殊的环境,尽管非洲大陆是人类的起源,但在非洲尚未有灭绝的古人类基因组序列的报道。manbet手机版因此,古人类对今天非洲人的基因变异的贡献仍然不太清楚。manbet手机版然而,在没有可用的参考古基因组的情况下,已经推断出古祖先的片段,并提出在西非人口中存在“幽灵”古祖先[52]。manbet手机版在非洲个体中也发现了尼安德特人的祖先,这一发现可以通过两种可能的解释:一是从欧洲祖先迁移回非洲,二是在尼安德特人离开非洲之前,他们的基因流入了尼安德特人。manbet手机版据称,尼安德特人的类似序列实际上可以在每一个当代现代人类基因组中被识别出来,分析到目前为止[53]。
manbet手机版现存最大的谜团之一是,为什么现代人能如此成功地进行扩张,以及为什么尼安德特人和丹尼索瓦人在适应欧亚环境几十万年后灭绝。manbet手机版观察到这些种群数量很少,而且近亲繁殖的程度相对较高,这给我们提供了一个线索,即他们可能无法与在种群规模上迅速扩张的现代人竞争。
manbet手机版与人类生理学的相关性
manbet手机版Svante Pääbo的工作提供了令人兴奋的可能性,可以在当今人类中寻找古老的基因变异,并研究它们与表型的关系。manbet手机版由于古人类在基因上已经适应了现代人类迁移到欧亚大陆的一些环境,来自尼安德特人和丹尼索瓦人的渗透事件可以引入等位基因,在他们离开非洲后,这些等位基因对他们在新环境中的生存能力产生了积极的影响。manbet手机版经过正选择的渐渗序列可以潜在地提供我们基因组部分的功能信息,这与所有活着的人类都有普遍关联。
manbet手机版来自尼安德特人的片段可以在人类基因组的大多数区域找到,尽管在基因组的不同部分频率不同。manbet手机版现在有有力的证据表明,这些信号中的一些代表了由渐渗事件衍生的古老的单倍型。manbet手机版一个明显的例子是丹尼索瓦人缺氧途径基因EPAS1的衍生版本,它赋予了在高海拔生存的遗传优势,在今天的藏人[54]中发现了它。manbet手机版跨越EPAS1基因的基因组区域包含一组snp,这些snp在丹尼索瓦人和藏族人的基因组中都有发现,在汉族人中也有非常低的频率。manbet手机版单倍型特异性snp的边界定义了一个32.7 kb的区域,这个长度与丹尼索瓦人和丹尼索瓦人时期发生的渗透事件一致manbet手机版智人manbet手机版共存。manbet手机版这种单倍型结构在全球任何其他种群中都没有观察到。
manbet手机版另一个有趣的基因渗入的例子是一组编码三个toll样受体的基因,manbet手机版TLR6-TLR1-TLR10manbet手机版,已知参与微生物识别和过敏反应[55]。manbet手机版珍妮特·凯尔索及其合作者证明,当今人类携带三种不同的跨越这些基因的古代单倍型,这表明在我们与古代亲戚的互动历史中发生了几次涉及这些基因的基因渗透事件。manbet手机版这些单倍型在目前的人类群体中出现的频率是可变的,但惊人的高,这表明它们提供了良好的生物学效应。manbet手机版其他的基因渗透事件的例子包括编码抗病毒信号分子Stat2的单倍型,在美拉尼西亚人[56]中出现频率很高,以及涉及2 ' -5 ' -寡聚腺苷酸合成酶(OAS)基因簇的单倍型,该基因簇也在抗病毒信号传递中发挥作用,从欧亚人[57]的一个亚群中从尼安德特人遗传而来。manbet手机版Pääbo和Zeberg最近的研究表明,染色体3和12上的古老等位基因(后者涉及OAS)可以影响SARS-CoV-2感染期间呼吸衰竭的风险[58,59]。manbet手机版这些和其他研究表明,关于基因导入对我们生理机能的功能影响,还有很多有待了解。
manbet手机版David Reich的团队与Pääbo合作,系统地推断了人类基因组中尼安德特人祖先的程度,并将一组已识别的等位基因与全基因组关联研究中识别的snp交叉,从而与一系列与疾病相关的表型[46]相关联。manbet手机版他们还发现,如Joshua Akey[48]小组所发现的那样,拥有高频率的尼安德特人等位基因的区域对影响角蛋白丝的基因来说是丰富的。manbet手机版英国生物银行(UK biobank)等数据库已经收集了大量的表型和基因型,包括可能从古人类基因中渗入的变体。manbet手机版利用这一资源,尼安德特人衍生的变异也与非疾病表型相关,如现代欧洲人[60]的色素沉着和睡眠模式。
manbet手机版另外还有一些例子表明,古等位基因与人类疾病和非疾病表型之间存在关联。manbet手机版当与我们的远古亲戚混合时,现代人类的祖先遇到了一个已经适应的古老等位基因库,随着环境的变化,这些等位基因可能会有用。manbet手机版于是,古代人的基因流入manbet手机版智人manbet手机版对人类生理的影响已经发生,为阐明特定基因变异如何在分子水平上调节生物过程提供了令人兴奋的可能性。
manbet手机版如果没有Svante Pääbo和他的团队的坚持不懈的工作,这一见解是不可能实现的,他们不仅定义了尼安德特人的核基因组,而且意外地发现了一种完全未知的古人类,并对其核基因组进行了完整的描述。
manbet手机版是什么让我们成为独一无二的人类?
manbet手机版智人manbet手机版人类文化、社会结构和沟通能力的复杂性在动物物种中脱颖而出。manbet手机版可以认为,这种独特的“人性”源于现代人类谱系基因组的变化。manbet手机版获取古代基因组提供了令人兴奋的新可能性,可以识别将我们与古代古人类区分开来的关键遗传特征。manbet手机版作为人类最亲近的进化亲缘,它们的基因组为阐明人类生理和医学相关的特定特征提供了参考点(manbet手机版图4manbet手机版).
manbet手机版在基因组中大约有31000个单核苷酸的位置,来自世界各地的现代人只携带一个新的(衍生的)核苷酸,而尼安德特人和丹尼索瓦人的基因组都携带从黑猩猩分裂出来以来保存下来的祖先核苷酸。manbet手机版在现今人类体内固定的变异中,约3000种位于调控区域,约30种影响假定的剪接位点,约100种影响蛋白质编码区域,改变氨基酸组成[61]。manbet手机版阐明独特的人类遗传变异的功能相关性是一个令人兴奋的挑战,与所有相关manbet手机版智人manbet手机版这是Svante Pääbo目前的主要研究方向之一。

manbet手机版图4。manbet手机版古代基因组为识别现代人与尼安德特人和丹尼索瓦人的关键基因特征提供了令人兴奋的新可能性。
manbet手机版卡洛琳斯卡学院教授古尼拉·卡尔松·赫斯塔姆博士(manbet手机版(电子邮件保护)manbet手机版)
manbet手机版诺贝尔委员会成员
manbet手机版安娜·韦德尔,医学博士,瑞典卡罗林斯卡学院教授manbet手机版(电子邮件保护)manbet手机版)
manbet手机版诺贝尔委员会成员
manbet手机版说明:这张Karlen
manbet手机版由卡罗林斯卡学院的50名教授组成的诺贝尔大会颁发诺贝尔生理学或医学奖。狗万世界杯manbet手机版诺贝尔委员会对提名进行评估。manbet手机版自1901年以来,诺贝狗万世界杯尔奖一直授予那些为人类利益做出最重要发现的科学家。
manbet手机版诺贝尔奖是诺贝尔基金会的注册商标
manbet手机版参考文献
- manbet手机版坎恩,r.l.,斯通金和A.C.威尔逊,manbet手机版线粒体DNA与人类进化。manbet手机版自然,1987年。manbet手机版325manbet手机版(6099): 31-6页。
- manbet手机版Lander, e.s.,等人,manbet手机版人类基因组的初步测序和分析。manbet手机版自然,2001年。manbet手机版409manbet手机版(6822): 860 - 921页。
- manbet手机版文特尔,j.c.等人,manbet手机版人类基因组的序列。manbet手机版科学,2001。manbet手机版291manbet手机版(5507): 1304 - 51页。
- manbet手机版Nurk, S.等人,manbet手机版人类基因组的完整序列。manbet手机版科学,2022。manbet手机版376manbet手机版(6588): 44-53页。
- manbet手机版基因组计划,C.等,manbet手机版从人口规模测序得出的人类基因组变异图。manbet手机版自然,2010年。manbet手机版467manbet手机版(7319): 1061 - 73页。
- manbet手机版马利克,S.等人,manbet手机版西蒙斯基因组多样性项目:来自142个不同人群的300个基因组。manbet手机版自然,2016年。manbet手机版538manbet手机版(7624): 201 - 206页。
- manbet手机版斋木,r.k.,等人,manbet手机版β -珠蛋白基因组序列的酶促扩增和酶切位点分析诊断镰状细胞性贫血。manbet手机版科学,1985。manbet手机版230manbet手机版(4732): 1350 - 4页。
- manbet手机版Paabo和A.C. Wilson,manbet手机版聚合酶链反应揭示克隆产物。manbet手机版自然,1988年。manbet手机版334manbet手机版(6181): 387 - 8页。
- manbet手机版布里格斯,a.w.等人,manbet手机版尼安德特人基因组DNA序列的损伤模式。manbet手机版美国国家科学院学报,2007。manbet手机版104manbet手机版(37): 14616 - 21页。
- manbet手机版Hoss, M.等人,manbet手机版古组织DNA损伤与DNA序列检索。manbet手机版核酸学报,1996。manbet手机版24manbet手机版(7): 1304 - 7页。
- manbet手机版Paabo、S。manbet手机版古代DNA:提取、表征、分子克隆和酶扩增。manbet手机版美国科学院学报,1989。manbet手机版86manbet手机版(6): 1939 - 43页。
- manbet手机版Hoss M. S. Paabo,manbet手机版用硅基纯化法从更新世骨骼中提取DNA。manbet手机版核酸学报,1993。manbet手机版21manbet手机版(16): 3913 - 4页。
- manbet手机版Poinar, H.N,等人,manbet手机版氨基酸外消旋化与古DNA的保存。manbet手机版科学,1996。manbet手机版272manbet手机版(5263): 864 - 6页。
- manbet手机版林达尔T。manbet手机版DNA初级结构的不稳定和衰变。manbet手机版自然,1993年。manbet手机版362manbet手机版(6422): 709 - 15页。
- manbet手机版van der Valk, T.等人,manbet手机版数百万年前的DNA揭示了猛犸象的基因组历史。manbet手机版自然,2021年。manbet手机版591manbet手机版(7849): 265 - 269页。
- manbet手机版Krings, M.等人,manbet手机版尼安德特人的DNA序列和现代人的起源。manbet手机版细胞,1997。manbet手机版90manbet手机版(1): 19-30页。
- manbet手机版Ovchinnikov,静脉注射等,manbet手机版对北高加索尼安德特人DNA的分子分析。manbet手机版自然,2000年。manbet手机版404manbet手机版(6777): 490 - 3页。
- manbet手机版Krings, M.等人,manbet手机版尼安德特人基因多样性的观点。manbet手机版Nat麝猫,2000年。manbet手机版26manbet手机版(2): 144 - 6页。
- manbet手机版Serre, D.等人,manbet手机版没有证据表明尼安德特人的mtDNA对早期现代人类有贡献。manbet手机版公共科学图书馆杂志,2004年。manbet手机版2manbet手机版(3): E57页。
- manbet手机版马古利斯,M.等人,manbet手机版微制造高密度皮升反应器的基因组测序。manbet手机版自然,2005年。manbet手机版437manbet手机版(7057): 376 - 80页。
- manbet手机版格林,r.e.等,manbet手机版对尼安德特人DNA的100万个碱基对进行分析。manbet手机版自然,2006年。manbet手机版444manbet手机版(7117): 330 - 6页。
- manbet手机版努南,J.P,等人,manbet手机版尼安德特人基因组DNA的测序和分析。manbet手机版科学,2006。manbet手机版314manbet手机版(5802): 1113 - 8页。
- manbet手机版沃尔,J.D.和S.K.金,manbet手机版尼安德特人基因组DNA序列的不一致。manbet手机版公共科学图书馆麝猫,2007年。manbet手机版3.manbet手机版(10): 1862 - 6页。
- manbet手机版Maricic T. S. Paabo,manbet手机版对从少量DNA制备的454测序文库进行优化,允许对两个DNA链进行序列测定。manbet手机版生物学技术,2009年。manbet手机版46manbet手机版(1)页51-2,54-7。
- manbet手机版Kircher M., U. Stenzel和J. Kelsomanbet手机版利用机器学习策略改进了Illumina基因组分析仪的基调用。manbet手机版基因组医学杂志,2009年。manbet手机版10manbet手机版(8): R83页。
- manbet手机版格林,r.e.等,manbet手机版通过高通量测序确定的完整尼安德特人线粒体基因组序列。manbet手机版细胞,2008。manbet手机版134manbet手机版(3): 416 - 26页。
- manbet手机版格林,r.e.等,manbet手机版尼安德特人基因组序列草图。manbet手机版科学,2010。manbet手机版328manbet手机版(5979): 710 - 722页。
- manbet手机版Krause, J.等人,manbet手机版来自南西伯利亚的一种未知古人类的完整线粒体DNA基因组。manbet手机版自然,2010年。manbet手机版464manbet手机版(7290): 894 - 7页。
- manbet手机版Reich, D,等人,manbet手机版西伯利亚丹尼索瓦洞穴一个古人类群的遗传历史。manbet手机版自然,2010年。manbet手机版468manbet手机版(7327): 1053 - 60页。
- manbet手机版梅耶,M.等人,manbet手机版一个古老丹尼索瓦人的高覆盖率基因组序列。manbet手机版科学,2012。manbet手机版338manbet手机版(6104): 222 - 6页。
- manbet手机版普鲁弗,K.等人,manbet手机版来自阿尔泰山脉的尼安德特人的完整基因组序列。manbet手机版自然,2014年。manbet手机版505manbet手机版(7481): 43-9页。
- manbet手机版Castellano, S.等人,manbet手机版三个尼安德特人完整外显子的编码变异模式。manbet手机版美国科学院学报,2014。manbet手机版111manbet手机版(18): 6666 - 71页。
- manbet手机版普鲁弗,K.等人,manbet手机版克罗地亚Vindija洞穴的高覆盖率尼安德特人基因组。manbet手机版科学,2017。manbet手机版358manbet手机版(6363): 655 - 658页。
- manbet手机版Hajdinjak, M.等人,manbet手机版重建晚期尼安德特人的基因历史。manbet手机版自然,2018年。manbet手机版555manbet手机版(7698): 652 - 656页。
- manbet手机版索耶,S.等人,manbet手机版两个丹尼索瓦人的细胞核和线粒体DNA序列。manbet手机版美国科学院学报,2015。manbet手机版112manbet手机版(51): 15696 - 700页。
- manbet手机版Slon, V.等人,manbet手机版第四个丹尼索瓦人。manbet手机版2017年Sci副词。manbet手机版3.manbet手机版(7): p . e1700186。
- manbet手机版Slon, V.等人,manbet手机版尼安德特人的母亲和丹尼索瓦人的父亲的后代的基因组。manbet手机版自然,2018年。manbet手机版561manbet手机版(7721): 113 - 116页。
- manbet手机版Mafessoni, F.等人,manbet手机版Chagyrskaya Cave的高覆盖率尼安德特人基因组。manbet手机版美国国家科学院,2020。manbet手机版117manbet手机版(26): 15132 - 15136页。
- manbet手机版梅耶,M.等人,manbet手机版中更新世司马德洛斯胡索古人类的核DNA序列。manbet手机版自然,2016年。manbet手机版531manbet手机版(7595): 504 - 7页。
- manbet手机版Rasmussen, M.等人,manbet手机版一个灭绝的古爱斯基摩人的古人类基因组序列。manbet手机版自然,2010年。manbet手机版463manbet手机版(7282): 757 - 62页。
- manbet手机版傅,Q,等,manbet手机版来自西西伯利亚的四万五千年前现代人的基因组序列。manbet手机版自然,2014年。manbet手机版514manbet手机版(7523): 445 - 9页。
- manbet手机版傅,Q,等,manbet手机版来自罗马尼亚的早期现代人,其祖先是尼安德特人。manbet手机版自然,2015年。manbet手机版524manbet手机版(7564): 216 - 9页。
- manbet手机版Slon, V.等人,manbet手机版更新世沉积物中的尼安德特人和丹尼索瓦人DNA。manbet手机版科学,2017。manbet手机版356manbet手机版(6338): 605 - 608页。
- manbet手机版扎瓦拉,我,等人,manbet手机版更新世沉积物DNA揭示了丹尼索瓦洞穴古人类和动物群的更替。manbet手机版自然,2021年。manbet手机版595manbet手机版(7867): 399 - 403页。
- manbet手机版Vernot, B.等人,manbet手机版利用洞穴沉积物中的细胞核和线粒体DNA挖掘尼安德特人的种群历史。manbet手机版科学,2021。manbet手机版372manbet手机版(6542)。
- manbet手机版Sankararaman, S.等人,manbet手机版现代人的尼安德特人祖先的基因组图谱。manbet手机版自然,2014年。manbet手机版507manbet手机版(7492): 354 - 7页。
- manbet手机版谢弗,北卡罗莱纳州,b·夏皮罗和r·e·格林,manbet手机版人类、尼安德特人和丹尼索瓦人基因组的祖先重组图。manbet手机版2021年Sci副词。manbet手机版7manbet手机版(29)。
- manbet手机版Vernot, b和j。m。Akey,manbet手机版从现代人类基因组中复活幸存的尼安德特人血统。manbet手机版科学,2014。manbet手机版343manbet手机版(6174): 1017 - 21页。
- manbet手机版Reich, D,等人,manbet手机版丹尼索瓦混血儿和第一批现代人类扩散到东南亚和大洋洲。manbet手机版Am J Hum Genet, 2011。manbet手机版89manbet手机版(4): 516 - 28页。
- manbet手机版维拉尼亚,F.A., J.G. Schraiber,manbet手机版尼安德特人和现代人之间的多次杂交。manbet手机版Nat Ecol Evol, 2019。manbet手机版3.manbet手机版(1): 39-44页。
- manbet手机版Choin, J.,等人,manbet手机版基因组对大洋洲种群历史和生物适应的洞察。manbet手机版自然,2021年。manbet手机版592manbet手机版(7855): 583 - 589页。
- manbet手机版Durvasula, A.和S. Sankararaman,manbet手机版在非洲人口中恢复幽灵古代人渗入的信号。manbet手机版2020年Sci副词。manbet手机版6manbet手机版(7): p . eaax5097。
- manbet手机版陈,L.,等人,manbet手机版在非洲个体中识别和解释明显的尼安德特人祖先。manbet手机版细胞,2020。manbet手机版180manbet手机版(4): p. 677-687 e16。
- manbet手机版Huerta-Sanchez, E.等人manbet手机版藏人的海拔适应是由丹尼索瓦人样DNA的渗入引起的。manbet手机版自然,2014年。manbet手机版512manbet手机版(7513): 194 - 7页。
- manbet手机版Dannemann, M,点manbet手机版安德烈斯和J.凯尔索,manbet手机版尼安德特人和丹尼索瓦人样单倍型的渗入有助于人类toll样受体的适应性变异。manbet手机版Am J Hum Genet, 2016。manbet手机版98manbet手机版(1): 22-33页。
- manbet手机版门德斯,f.l., J.C.沃特金斯和M.F.汉默,manbet手机版STAT2的单倍型由尼安德特人渗入,在巴布亚新几内亚作为积极选择的候选。manbet手机版Am J Hum Genet, 2012。manbet手机版91manbet手机版(2): 265 - 74页。
- manbet手机版门德斯,f.l., J.C.沃特金斯和M.F.汉默,manbet手机版尼安德特人OAS免疫基因簇遗传变异的起源。manbet手机版Mol Biol Evol, 2013。manbet手机版30.manbet手机版(4): 798 - 801页。
- manbet手机版泽伯格,H.和S.帕博,manbet手机版严重COVID-19的主要遗传风险因素来自尼安德特人。manbet手机版自然,2020年。manbet手机版587manbet手机版(7835): 610 - 612页。
- manbet手机版泽伯格,H.和S.帕博,manbet手机版一个与预防严重COVID-19相关的基因组区域继承自尼安德特人。manbet手机版美国国家科学院学报,2021年。manbet手机版118manbet手机版(9)。
- manbet手机版丹内曼,M. J.凯尔索,manbet手机版尼安德特人对现代人类表型变异的贡献。manbet手机版Am J Hum Genet, 2017。manbet手机版101manbet手机版(4): 578 - 589页。
- manbet手机版Paabo、S。manbet手机版人类状况——分子方法。manbet手机版细胞,2014。manbet手机版157manbet手机版(1): 216 - 26页。
manbet手机版观看2022年诺贝尔奖公告
manbet手机版即将到来的
manbet手机版观看公告的直播。

